
Lerntext Organellen
Roland Heynkes 22.3.2025, CC BY-SA-4.0 DE
If you cannot read German-language texts, you could try a translation into your native language with DeepL.
Dieser Schulbuch-unabhängige Selbstlern-Hypertext soll es Schülern (m,w,d) ermöglichen, selbständig und im eigenen Tempo zu lernen oder zu wiederholen, was gebildete Menschen über Organellen wissen sollten und was in der Qualiphase der gymnasialen Oberstufe als Vorwissen erwartet wird.
In Anlehnung an die Organe vielzelliger Lebewesen bezeichnet man eine vom Rest der eukaryotischen Zelle abgegrenzte Struktur mit einer bestimmten Aufgabe als Organell oder Organelle (Mehrzahl = Organellen). Organellen sind also strukturelle und funktionale Einheiten. Ähnlich wie die Organe in einem Organismus arbeiten in eukaryotischen Zellen Organellen räumlich und zeitlich koordiniert zusammen. Organellen grenzen sich vom Zytoplasma ab, um in ihren Funktionen nicht von anderen Teilen der Zelle gestört zu werden oder selbst der Zelle zu schaden. Normalerweise geschieht diese Kompartimentierung genannte Abgrenzung durch eine Membran, welche das Organell umgibt. Viele Biologen meinen, solche strukturellen und funktionellen Einheiten müssten von einer Membran umgeben sein, um Organell genannt werden zu können. Typische Organellen nach dieser engen Definition sind Zellkern, endoplasmatisches Retikulum, Golgi-Apparat, Endosomen, Lysosomen, Mitochondrien und Chloroplasten. Weniger starr an veralteten Definitionen hängende Biologen zählen zu den Organellen auch strukturell vom Cytoplasma abgegrenzte funktionelle Einheiten, bei denen die Abgrenzung nicht durch eine Membran erfolgt. Beispiele dafür sind die Proteasomen oder Augenfleck und Zellmund in Einzellern. Das Biologiebuch "Natura Oberstufe" beschreibt auf einer Doppelseite über Bau und Funktionen von Zellorganellen auch Mikrotubuli, Mikrofilamente, Centriolen und die Zellwand der Pflanzenzelle und zählt sogar die Ribosomen explizit zu den Organellen. Mir persönlich geht das allerdings zu weit, denn dann gäbe es Organellen (Ribosomen) in Organellen (Mitochondrien und Chloroplasten) und man müsste sich fragen, warum nicht auch die DNA- und RNA-Polymerasen Organellen sind. Organellen sind nicht einfach zufällig in der Zelle verteilt, sondern werden von Mikrotubuli an bestimmten Stellen gehalten oder mit Motorproteinen bewegt. Insgesamt ist die Fläche der Organellenmembranen einer Zelle viel größer als die der Zellmembran.
Im Gegensatz zu den einfacher aufgebauten Prokaryoten besitzt bei den komplexeren Eukaryoten jede noch teilungsfähige Zelle einen Zellkern. Dessen aus zwei Membranen aufgebaute Kernhülle umschließt und schützt den größten Teil des Erbmaterials einer Zelle. Für einen geregelten Informationsaustausch zwischen dem Zellkern und dem Rest der Zelle gibt es in der Kernhülle große Kernporen.
 |
| Woland Messer, GNU Free Documentation License |
|
Das folgende Schema zeigt zusätzlich Elemente des Zytoskeletts, wenn auch nicht im Zusammenhang. Die großen schwarzen Öffnungen in der Zellmembran stellen Vesikel dar, die gerade mit der Zellmembran verschmelzen (Exozytose) oder sich von ihr nach innen abschnüren (Endozytose).
| Schema einer tierischen Zelle |
|---|
 |
| Mariana Ruiz Villarreal, public domain |
Das folgende Modell einer tierischen Zelle im Museum des indischen Kolkata scheint die Vorlage der Zeichnung von Woland Messer zu sein.
| Modell einer tierischen Zelle im Museum des indischen Kolkata |
|---|
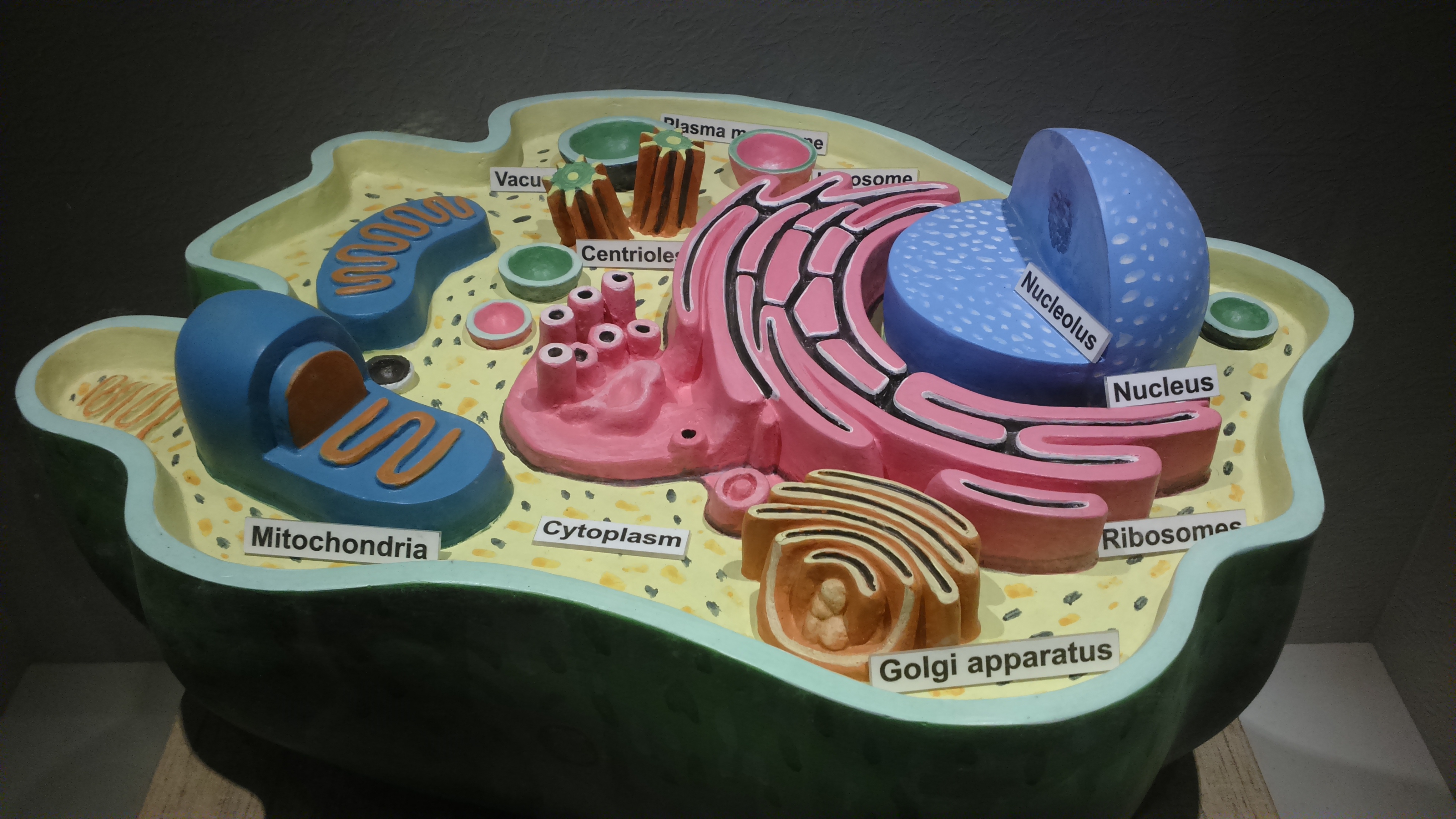 |
| anonym, CC BY-SA 4.0 |
Zur selbständigen Erarbeitung dieses Lerntext-Kapitels gibt es ein Lernmodul mit klausurähnlichen Aufgaben (PDF)
In normalen Eukaryoten-Zellen (tierischen, pflanzlichen oder pilzlichen) färben bestimmte Farbstoffe besonders den meistens großen und kugeligen Zellkern (Nukleus), den es in Prokaryoten (Bakterien und Archäen) nicht gibt. Das folgende Foto von einem dünnen Schnitt aus einer Zwiebelwurzelspitze zeigt die großen Zellkerne junger Pflanzen-Zellen.
Im Inneren des Zellkerns befindet sich die Karyolymphe. In ihr schwimmen das Chromatin und meistens außerdem die weniger stark angefärbten Nukleoli, die in einem eigenen Kapitel beschrieben werden. Chromatin nennt man die Gesamtheit der in einem Zellkern besonders intensiv anfärbbaren DNA-Protein-Komplexe. Der Name Chromosom wurde zusammengesetzt aus den griechischen Begriffen chroma für Farbe und soma für Körper. Wichtig ist aber nicht die Färbbarkeit der Chromosomen, sondern ihre Funktion als Rezeptbücher des Lebens.
| gefärbte Zellkerne in jungen Pflanzen-Zellen |
|---|
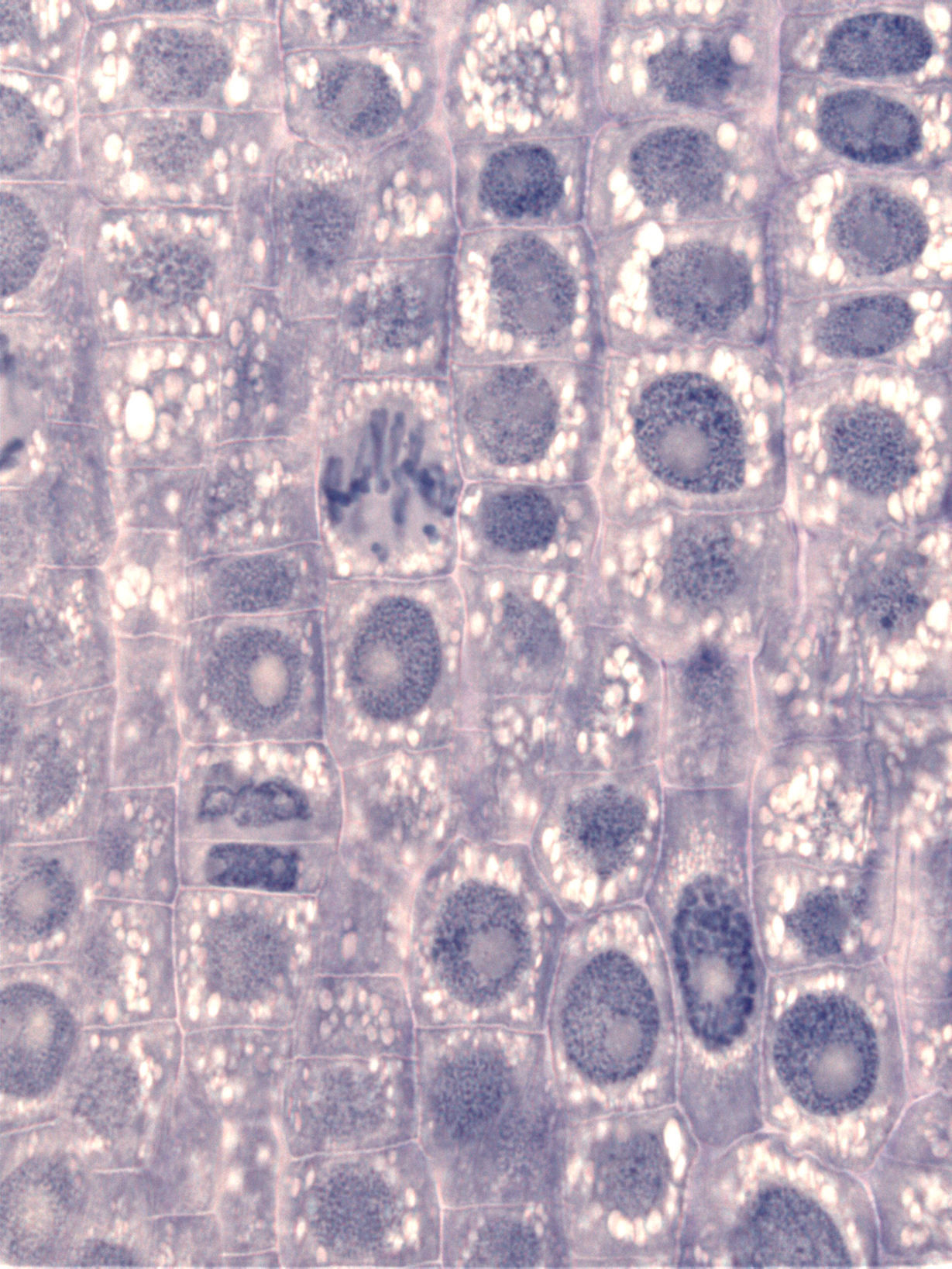 |
| Dr. phil.nat Thomas Geier, CC BY-SA 3.0 |
Der Zellkern enthält und beschützt vor allem die beiden von Vater und Mutter geerbten Baupläne jeder noch lebensfähigen Zelle, die zusammen Genom genannt werden. Im Prinzip enthalten alle Zellkerne eines eukaryotischen Lebewesens die gleichen Erbinformationen, die schon der Zellkern der Zygote enthielt. Ausnahmen sind die Geschlechtszellen sowie Lymphozyten, bei deren Reifung Gene für B- oder T-Zell-Rezeptoren oder Antikörper individuell zusammengesetzt werden. Nicht ganz vergessen darf man natürlich auch die Mutationen, die sich bei jeder Zellteilung einschleichen und dafür sorgen, dass kaum zwei Zellen eines vielzelligen Organismus wirklich identisch sind. Solche minimalen Unterschiede sind aber viel zu gering und zufällig, um die Unterschiede zwischen den über 200 Zelltypen des Menschen zu erklären. Der Zellkern kann auch nicht selber aus sich heraus erkennen, welche Bauanleitungen (Gene) die Zelle aktivieren muss. Denn es gibt neben dem Genom nichts anderes in einem Zellkern, was als Kommandozentrale fungieren könnte. Der Zellkern kann also nicht dafür verantwortlich sein, dass Nervenzellen und Muskelzellen so unterschiedlich sind. Darum wäre es falsch und irreführend, den Zellkern mit einer Kommandozentrale oder dem Rathaus einer Stadt zu vergleichen. Nicht er, sondern die ganze Zelle entscheidet vor allem aufgrund vielfältiger Signale von anderen Zellen, welche der vielen Rezepte (Gene) für einzelne Eiweiße oder RNAs sie gerade benötigt.
Die Funktion des Zellkerns entspricht vielmehr einer Bibliothek mit sehr wertvollen, über Generationen vererbten Kochbüchern. In menschlichen Zellen enthält diese Bibliothek Zellkern Zigtausende Baupläne für Proteine und RNAs, die unseren Zellen als Bausteine und Werkzeuge dienen. Selbstverständlich werden in einer Bibliothek niemals alle Bücher gleichzeitig gelesen. Genauso benutzt keine Zelle alle ihre Gene genannten Baupläne. Aber wenn eine Zelle ein bestimmtes Rezept (Gen) braucht, dann schickt sie Signale in den Zellkern. Dadurch werden die gerade benötigten Gene aktiviert.
Damit sie nicht beschädigt werden und jederzeit verfügbar sind, darf man die unersetzlichen Bücher einer Bibliothek nicht ausleihen, sondern nur fotokopieren. Die Bücher des Zellkerns heißen Chromosomen und auch sie müssen zwischen den Kernteilungen immer im Zellkern bleiben. Dort werden Kopien von den gerade benötigten Genen gemacht. Der allgemeinverständliche Begriff Kopie trifft die Sache allerdings nicht ganz naturwissenschaftlich exakt. Denn während die Gene in den Chromosomen aus dem Material DNA (Desoxyribonukleinsäure) bestehen, werden die "Kopien" aus RNA (Ribonukleinsäure) hergestellt. Genau genommen sind deshalb die RNAs keine Kopien, sondern Transkripte der DNA. Darum sprechen Biologen auch nicht von Kopieren, sondern nennen den Vorgang Transkription. Aber entscheidend für die in ihm steckende Information ist nicht das Material, sondern die Reihenfolge (Sequenz) seiner Grundbausteine (Monomere), die man bei DNA und RNA Nukleotide nennt.
Den Zellkern verlassen durch die Kernporen nur Transkripte. Weil sie wie Boten (englisch: messenger) eine Botschaft (englisch: message) aus dem Zellkern ins Cytoplasma bringen, bezeichnet man diese RNAs als Boten-RNS oder englisch messenger-RNAs. Es gibt aber auch viele Gene, deren Transkripte keine mRNAs sind, sondern beispielsweise direkt als ribosomale RNAs (rRNAs) oder tRNAs genutzt werden. In der Cytoplasma genannten, Gel-artigen Grundsubstanz der Zelle werden die mRNAs von Ribosomen gebunden. An ihnen werden die Nukleotidsequenzen der mRNAs in die Aminosäuresequenzn von Proteinen (Eiweißen) übersetzt. Dieser Vorgang heißt Translation (Übersetzung). Darum sind die Gene Rezepte für die Proteinbiosynthese.
|
| Gene codieren Proteine |
|---|
 |
Eukaryotische Zellen enthalten zusätzliche Erbinformationen in Mitochondrien und bei pflanzlichen Zellen außerdem in Chloroplasten. Mitochondrien und Chloroplasten enthalten ihre eigenen kleinen Baupläne, weil sie wie winzige Bakterien in eukaryotischen Zellen leben und sich selbständig durch Zellteilung vermehren. Insofern ist schon jede einzelne eukaryotische Zelle ganz ähnlich wie eine Flechte eine Symbiose (genauer: Endosymbiose) aus einer großen schützenden Zelle und ihren lebenswichtige Energie liefernden Endosymbionten. Wegen dieser relativen Unabhängigkeit (Semiautonomie) der Mitochondrien und Chloroplasten werden ihre Baupläne oft vergessen, wenn von der Genom genannten Gesamtheit aller Erbinformationen eines eukaryotischen Lebewesens die Rede ist. Das ist aber nicht korrekt, sofern man beispielsweise uns Menschen als Lebewesen und nicht nur als Symbiose zwischen Mensch und Mitochondrien betrachtet. Weitere Informationen zur Endosymbionten-Theorie findet man im Lerntext Endosymbionten-Theorie.
Der Zellkern ist keine Steuerungszentrale, sondern wie eine Bibliothek mit wertvollen Kochbüchern. Die Kochbücher im Zellkern heißen Chromosomen und enthalten viele Rezepte, die wir Gene nennen. Diese Gene sind Bauanleitungen für Proteine und spezielle RNAs, die selber wie die Proteine als Werkzeuge oder Bausteine wirken können. Wie die Rezepte eines Kochbuchs werden auch die Gene in den Chromosomen nie alle gleichzeitig gebraucht. Jeder Zelltyp aktiviert je nach Entwicklungsphase und Situation eine andere Menge von Genen. So produziert jede Zelle immer nur die Werkzeuge und Bausteine, die sie tatsächlich gerade braucht. Wenn eine Tochterzelle einer Stammzelle sich schrittweise immer mehr zu einer hochspezialisierten (voll differenzierten) Zelle entwickelt, gibt es immer mehr Gene, die sie nie wieder benötigen wird. Darum fragten sich Wissenschaftler in den 1950er Jahren, ob sich differenzierende Zellen ihre überflüssig gewordenen Gene verlieren oder unwiederbringlich inaktivieren. Eines von vielen Experimenten zu dieser Frage soll das folgende Schema zeigen. Leider konnte ich die Veröffentlichung mit genau dieser Variante des Experiments noch nicht finden, aber Sir John Bertrand Gurdon erwähnte nebenbei anlässlich der Verleihung seines Nobelpreises, dass er zeitweise auch Albinofrösche als Zellkern-Spender verwendete, weil sein sonst mit anderen Markern durchgeführtes Experiment dadurch auch für Laien besonders überzeugend wurde (J. B. Gurdon - The egg and the nucleus: a battle for supremacy - Development 2013 140: 2449-2456). Wer sich genauer mit den Gurdon-Experimenten beschäftigen möchte, findet Details zu etlichen seiner unzähligen Experimente im Lerntext Xenopus-Nukleus-Transplantationen.
| Xenopus-Zellkern-Transfer-Experiment von Sir John Bertrand Gurdon |
|---|
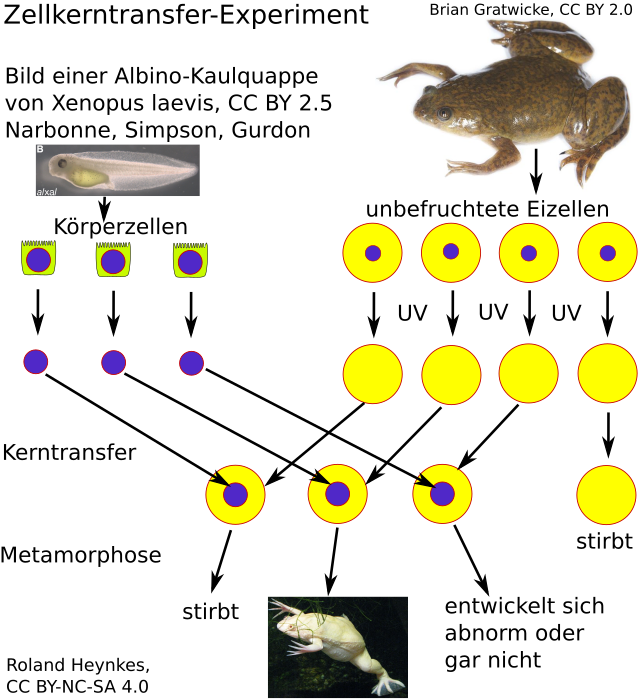
|
| In den frühen 1960er Jahren führte der später dafür mit dem Nobelpreis ausgezeichnete John Gurdon Zellkern-Transfer-Experimente mit südafrikanischen Krallenfröschen durch. Er zerstörte die DNA haploider Zellkerne unbefruchteter Eizellen normaler Krallenfrösche mit UV-Strahlung. Dadurch fehlten diesen Empfängerzellen funktionsfähige Gene und sie wären abgestorben. Gleichzeitig enthielten aber diese Empfängerzellen im Cytoplasma und in ihren bestrahlten Zellkernen noch fast alle Proteine und viele andere für eine normale Embryonal-Entwicklung nötige Faktoren. Gurdon injizierte in diese nun praktisch Gen-freien Eizellen intakte Zellkerne aus bereits differenzierten, diploiden Körperzellen von Albino-Kaulquappen, die aufgrund einer Mutation den Farbstoff des Wildtyps nicht produzieren können. Aus manchen der so manipulierten Eizellen entwickelten sich Albino-Frösche. Damit zeigte er, dass sich die hierfür verantwortliche Erbinformation (Genom) im Zellkern und nicht im Rest der Zelle befindet. Vor allem zeigte er, dass nicht mehr benötigte Gene während der Differenzierung nicht verloren gehen, sondern lediglich inaktiviert werden. Er zeigte damit aber nicht, dass der Zellkern die Lebensvorgänge steuert. Ganz im Gegenteil zeigt dieses Experiment, dass die Eizelle den Zellkern umprogrammiert. Denn der hatte die Gene für eine differenziertere Körperzelle bereitgestellt und nun mussten in ihm die Gene für die Embryogenese reaktiviert werden. Das dies meistens so einfach nicht gelingt, liegt an dem komplexen Zusammenspiel epigenetisch vererbter Veränderungen des Zellkerns und seiner Regulation durch die Zelle sowie deren Regulation durch Nachbarzellen und Hormone. |
Das Lexikon der Biologie beschreibt die frühen Experimente mit Kerntransplantationen und übernimmt dabei eine damals (nicht von Gurdon) verbreitete Fehlinterpretation, diese Experimente hätten den "Nachweis von Steuerfunktionen des Kerns im Hinblick auf die Zelldifferenzierung" geliefert. In unzähligen Experimenten haben Gurdon und seine Arbeitsgruppe aber ganz im Gegenteil gezeigt, dass während ihrer Reifung die Geschlechtszellen die Benutzbarkeit der Gene in ihren Zellkernen epigenetisch festlegen und wichtige Steuerungsfaktoren in den Zellkernen und im Cytoplasma anhäufen, damit die Eizelle nach der Befruchtung durch ein Spermium die richtigen Gene zum genau richtigen Zeitpunkt für die passende Zeitdauer aktiviert und exakt die richtige Menge von Transkripten produziert.
Acetabularia heißen aus nur einer riesigen Zelle bestehende Schirmalgen. Aufgrund ihrer Größe eignen sie sich gut für Experimente. Der Lerntext Acetabularia-Experimente beschreibt Experimente, mit denen schon vor dem zweiten Weltkrieg untersucht wurde, wie eine Zelle ihren Zellkern benutzt.
Ein Nukleolus oder mehrere Nukleoli innerhalb des Zellkerns einer eukaryotischen Zelle sind die Orte, an denen die größeren und die kleineren Ribosomen-Untereinheiten aus ribosomalen RNAs (rRNAs) und Proteinen zusammen gesetzt werden. Das geschieht nicht einfach überall im Zellkern, sondern in den Umgebungen der Gene für ribosomale RNAs. An ihnen werden die ribosomalen RNAs (rRNA) synthetisiert (Transkription).
Es gibt keine Membran zur Abgrenzung des auch Kernkörperchen genannten Nucleolus vom Rest des Zellkerns. Erkennbar werden die Orte der Ribosomen-Produktion durch ihre starke Lichtbrechung und dadurch, dass Farbstoffe im Zellkern zwar immer Nucleoli und das Chromatin anfärben, aber nicht gleich intensiv. Die Ribosomen-Untereinheiten verlassen durch die großen Kernporen den Zellkern und im Cytoplasma finden jeweils eine große und eine kleine Untereinheit zu einem kompletten Ribosom zusammen.
Der Zellkern einer eukaryotischen Zelle wird vom ihn umgebenden Zytoplasma durch eine Kernhülle getrennt. Sie besteht aus zwei je 7,5 nm dicken Kernmembranen mit einem 20 nm breiten Spalt dazwischen. Die Zellkernhülle wird vom rauen endoplasmatischen Retikulum gebildet bleibt mit diesem verbunden.
In der Zellkernhülle gibt es große Kernporen für den kontrollierten Austausch von Stoffen, Makromolekülen und Ribosomen-Untereinheiten zwischen Karyolymphe und Zytoplasma. Die Kernporen sind kompliziert aufgebaute Kanäle mit 60-100 nm Durchmesser und langen Tentakeln.
Raues endoplasmatisches Retikulum (rER) heißt der auf seiner Oberfläche mit Ribosomen besetzte, innere (dem Zellkern aufliegende) Teil des endoplasmatischen Retikulums (ER). Es besteht aus einem Netzwerk röhrenförmiger Membranen mit Zisternen genannten, großen, abgeflachten Aussackungen und ist wie der Zellkern ein unverzichtbares Organell jeder eukaryotischen Zelle, die noch teilungsfähig ist oder wenigstens noch Eiweiße und Membranen produziert. Man kann es mit den Fabriken und Werkstätten einer Stadt vergleichen, denn es produziert ständig neue Membranen und aus dem Zytoplasma zu exportierende Proteine sowie auf Lipid- und Protein-Oberflächen komplexe Kohlenhydrate. In die Membran des rER integrierte Enzyme setzen aus Grundbausteinen Lipide zusammen, die dann in der Membran verbleiben und deren Fläche vergrößern. Die auf dem rauen ER sitzenden Ribosomen synthetisieren die Proteine in die Membran des endoplasmatischen Retikulums hinein oder durch die Membran des endoplasmatischen Retikulums hindurch in dessen Inneres (Lumen) hinein. Die Ribosomen sitzen allerdings nicht permanent auf dem rauen endoplasmatischen Retikulum, sondern docken erst an, nachdem sie während der im Cytoplasma gestarteten Translation die ersten Codons der mRNA in ein erstes, kurzes Signal-Peptid übersetzt haben, welches anzeigt, dass da gerade ein Membran- oder Exportprotein entsteht. Sobald ein Ribosom auf der Membran des rauen ER sitzt, synthetisiert es den Rest der Aminosäurekette durch eine sich zeitweise öffnende Membranpore direkt in die Membran oder in eine Zisterne des ER hinein. Dort bilden sich dann die Tertiärstrukturen der für die Ausscheidung aus der Zelle oder eine Funktion auf der Zellmembranaußenseite bestimmten Proteine. Im ER bilden sich Disulfidbrücken zwischen Cysteinseitenketten und durch die auf der Innenseite der ER-Membran verankerte Oligosaccharyltransferasen werden viele Proteine im ER auch kovalent über die Aminogruppe einer Asparginseitenkette mit vorgefertigten kurzen, verzweigten Oligosacchariden verknüpft. Auch Quartärstrukturen aus mehreren Polypeptiden bilden sich hier. Aus dem ER exportiert werden nur korrekt gefaltete Proteine, die außerdem an ihren Carboxytermini keine aus 4 Aminosäuren bestehende Rückhaltesequenz aufweisen. Nach jeder Zellkern-Teilung bildet das raue endoplasmatische Retikulum eine neue Zellkernhülle und bleibt mit der äußeren Zellkern-Membran verbunden.
 |
| Blausen Medical Communications, Inc., CC BY 3.0 |
Glattes endoplasmatisches Retikulum heißt der nur in einigen Zellen eindeutig erkennbare, keine Ribosomen tragende, multifunktionelle Anteil des endoplasmatischen Retikulums (ER) in eukaryotischen Zellen. Es besteht aus einem Netzwerk röhrenförmiger Membranen mit Zisternen genannten, großen, abgeflachten Aussackungen. Es dient unter anderem dem Transport wichtiger Stoffe innerhalb der Zelle, der Calciumspeicherung (Muskelzellen), der Steroidhormon-, Lipid- und Glycogen-Synthese sowie der Glycogen-Spaltung, der Entgiftung und dem Hormon-Transport. Vom glatten ER schnüren sich Vesikel ab, die danach zum Golgi-Apparat transportiert werden.
| Der Golgi-Apparat ist nur elektronenmikroskopisch erkennbar und befindet sich zwischen dem endoplasmatischen Retikulum und der Zellmembran. Der Golgi-Apparat besteht aus 4-6, bei manchen Einzellern bis zu mehreren Hundert Dictyosomen genannten Stapeln flacher Membransäckchen (Zisternen). Auf der konvexen, dem ER zugewandten cis-Seite verschmelzen vom ER abgeschnürte Vesikel mit den Ausläufern des Golgi-Apparates zunächst zu einem cis-Golgi-Netzwerk aus Vesikeln und Zisternen. Darin vereinigen sich die im ER produzierten biologischen Makromoleküle mit durch Abschnürung von Vesikeln von der Zelloberfläche zurückgeholten Proteinen. Von der cis-Seite können sich aber auch Vesikel abschnüren, die zum ER wandern. Die vom ER ab- und der Zellmembran zugewandte Seite ist konkav, löst sich zum sogenannten trans-Golgi-Netzwerk auf und verschwindet durch die Abschnürung von Golgi-Vesikeln, während die inneren Zisternen nachrücken. Die Zisternen zwischen den Golgi-Netzwerken werden Golgi-Stapel genannt. Auch innerhalb dieser Stapel scheinen die Zisternen in beide Richtungen Vesikel auszutauschen. | |
 |
|
| Magnus Manske, CC BY 3.0 | |
| Dictyosom nennen Biologen einen Stapel von 3-8, selten bis zu 30 flachen, Membran-umhüllten Räumen mit durchschnittlich 1 µm Durchmesser, die man Zisternen nennt. Alle Dictyosomen gemeinsam bilden den Golgi-Apparat. Sie modifizieren, adressieren und schicken Proteine und Lipide an ihre Bestimmungsorte in oder außerhalb der Zelle. Dictyosomen entsprechen also Poststationen, Logistikzentren wie DHL oder den Warenlagern von Versandhandels-Unternehmen wie Amazon. In den Dictyosomen werden unter anderem Sekrete kondensiert und verpackt, Glykolipide synthetisiert und Proteine an den Aminosäuren Serin und Threonin glykosiliert, an den Zuckerresten sulfatiert, von Signalpeptiden befreit und teilweise durch Spaltung aktiviert. | |
| Aufgaben zur selbständigen Erarbeitung des Lernstoffes | |
|---|---|
| 1 | Was und wie groß sind Mitochondrien? |
| 2 | Wieviele Mitochondrien leben in einer Zelle? |
| 3 | Woran kann man sie so gut erkennen? |
| 4 | Wie sind Mitochondrien aufgebaut? |
| 5 | Was sind ihre Aufgaben? |
| 6 | Was ist ATP und welcher Prozess bildet es? |
| 7 | Warum wohl enthalten Mitochondrien DNA? |
| 8 | Überlege, wie die im Nukleus codierten Proteine in die Mitochondrien kommen könnten! |
| Hier geht es zu den Lösungen. | |

Mariana Ruiz Villarreal , public domain |
Mitochondrien sind 1-10 µm kleine, runde oder längliche Körperchen. Je nach Größe und Energiebedarf findet man in Zellen 1.000 - 100.000 Mitochondrien. Typisch für Mitochondrien ist die massiv gefaltete innere Membran innerhalb der äußeren. Mitochondrien haben eine äußere und eine röhren- (Tubuli) oder kamm- bzw. lamellenförmig (Christae = kammförmige) zwecks Oberflächenvergrößerung stark gefaltete innere Membran. Beide Membranen sind 7,5 nm dick und mindestens 10 nm von einander entfernt. Zwischen den Membranen gibt es eine feinkörnige Grundsubstanz namens Matrix. Enzyme in ihrer Matrix katalysieren den Zitronensäurezyklus, den Abbau von Fettsäuren und den Umbau von Aminosäuren. ATP ist die chemische Speicherform für Energie in der Zellen und wird in Mitochondrien im Verlauf der Atmungskette an den ATP-Synthase-Komplexen gebildet. Proteine werden im Cytoplasma synthetisiert und dann durch die Membranen in die Mitochondrien transportiert. |
| Mitochondrium, Mitochondrion oder Kraftwerk der Zelle nennt man ein wichtiges Organell der Eukaryoten-Zelle. Nach der weitestgehend akzeptierten Endosymbiontentheorie handelt es sich bei den Mitochondrien um Nachfahren von Bakterien, die sich vor Milliarden Jahren von Beute oder Krankheitserregern einer Archäen-Spezies zu deren Endosymbionten entwickelten. Sie besitzen einen eigenen Bauplan, vermehren sich durch Zellteilung und werden nicht vom Vater, sondern nur von der Mutter auf ihre Kinder übertragen. Die heute anscheinend nur noch innerhalb eukaryotischer Zellen lebensfähigen Mitochondrien bekommen vom Archäen-Partner Nährstoffe und Schutz vor Fressfeinden und liefern dafür so große Mengen des universell verwendbaren Trägers chemische Energie namens ATP, dass verglichen Archäen und Bakterien riesige eukaryotische Zellen entstehen konnten. Das Bakterium war nämlich in der Lage, Sauerstoff für eine besonders effektive Verdauung von Nahrung zu nutzen. Inzwischen hat das Bakterium einen Teil seiner nicht mehr benötigten Gene verloren, weil es von der Zelle versorgt wird. | |
Lysosomen nennt man die Organellen, die in tierischen Zellen der enzymatischen Verdauung und dem Recycling alter Zellbestandteile und Nahrung dienen. Sie verdauen Membranen zu Lipiden, Proteine zu Aminosäuren und Nukleinsäuren zu Nukleotiden, die dann zur Energie-"Gewinnung" oder wieder zum Aufbau zelleigener Makromoleküle genutzt werden können.
Primäre Lysosomen entstehen als Verdauungsvesikel durch Abschnürung von mit Verdauungsenzymen gefüllten Vesikeln vom Golgi-Apparat. Sie verschmelzen mit Endosomen zu sekundären Lysosomen, in derem durch Protonenpumpen angesäurtem Inneren etwa 40 hydrolytische Enzyme abgeschirmt vom Cytoplasma die Inhalte der ehemaligen Endosomen abbauen.
Peroxisomen sind vom endoplasmatischen Retikulum abgeschnürte Vesikel und winzige Entgiftungs-Organellen eukaryotischer Zellen. Sie erzeugen und nutzen H2O2 für einen oxydativen Abbau von Lipiden und giftigen Stoffen wie Alkohol.
Endosomen nennt man in der Zellbiologie größere Membranbläschen (Vesikel), welche direkt oder indirekt durch Abschnürung (Endozytose) von der Zellmembran entstehen. Man könnte diese Organellen auch Verdauungsorgane der Zelle nennen, die Verdaubares von außerhalb der Zelle enthalten. Bei der Phagozytose spricht man unmittelbar nach der Abschnürung von der Zellmembran von einem Endosom, bei der Rezeptor-vermittelten Endozytose erst nach dem Verschmelzen (Fusion) mehrerer von ihren Clathrin-Hüllen befreiter Vesikel. Zu richtigen Verdauungsorganellen der Zelle werden Endosomen aber erst durch die Verschmelzung mit primären Lysosomen, die verschiedene Verdauungs-Enzyme enthalten. Bevor die jungen/frühen Endosomen mit primären Lysosom zu sekundären Lysosom fusionieren, sortieren sie einen Teil der eingeschlossenen Makromoleküle zur Wiederverwendung aus. Gelegentlich nennt man die indirekt durch Rezeptor-vermittelte Endozytose gebildeten Endosomen auch Rezeptosomen.
| Aufgaben zur selbständigen Erarbeitung des Lernstoffes | |
|---|---|
| 1 | Definiere Plastid, Chromoplast, Leukoplasten und Chloroplast! |
| 2 | Nenne die Art der Plastiden-Vermehrung und was sie dafür benötigen! |
| Hier geht es zu den Lösungen. | |
Plastid nennen Biologen jedes Organell, das es nur in Pflanzen, Algen und einigen tierischen Einzellern gibt. Im Gegensatz zu anderen Organellen wie dem Zellkern, dem endoplasmatischen Retikulum oder dem Golgi-Apparat werden Plastide nicht von der großen Zelle gebildet. Man geht heute davon aus, dass Pflanzenzellen durch eine Endosymbiose aus tierischen Zellen bzw. tierischen Einzellern mit Cyanobakterien entstanden sind. Deshalb besitzen die Plastiden wie die Mitochondrien eigene kleine Genome, eigene Ribosomen und eine eigene Plasmamembran und vermehren sich in den Zellen als Endosymbionten selbständig durch Zellteilung.
Das folgende Schema der pflanzlichen Plastiden zeigt deren vermutete Verwandtschaft und wie sie auseinander hervorgehen können.
| Schema der pflanzlichen Plastiden | |
|---|---|
 | |
| Mariana Ruiz Villarreal, public domain | |
In den Zellen reifer Früchte und vieler Blüten findet man Chromoplasten genannte, durch Carotionide gelb- bis orange gefärbte Körnchen. In den farblosen Zellen pflanzlicher Speicherorgane wie Samen, Knollen oder Wurzelstöcken findet man unauffällige Körnchen ohne Farbstoffe. Sie heißen Leukoplasten und sie bauen aus Zucker Reservestärke auf.
Die wichtigsten Plastide sind die Chloroplasten der Grünalgen und höheren Pflanzen, denn ihre Fotosynthese ist die Lebensgrundlage fast aller Pflanzen, Pilze und Tiere.
| Aufgaben zur selbständigen Erarbeitung des Lernstoffes | |
|---|---|
| 1 | Was sind Chloroplasten und was ist ihre Aufgabe? |
| 2 | Welche Farbstoffe sind für die Fotosynthese erforderlich und welche Eigenschaften haben sie? |
| 3 | Woran erkennt man, dass Chloroplasten Zucker produzieren? |
| 4 | Wann und warum sieht man die Carotinoide in den Blättern unserer Laubbäume? |
| 5 | Für welche beiden Prozesse benötigen Chloroplasten Licht? |
| Hier geht es zu den Lösungen. | |
Viele Pflanzenzellen enthalten Chloroplasten genannte Organellen. Sie geben den Pflanzen ihre grüne Farbe und in ihnen findet die Fotosynthese statt. Ohne die Fotosynthese könnte kein Mensch leben, denn sie nutzt die Energie des Sonnenlichts, um die für uns lebenswichtigen Moleküle Traubenzucker und Sauerstoff aus Wasser und Kohlenstoffdioxid zu produzieren. Wie die Mitochondrien besitzen auch die Chloroplasten einen eigenen Bauplan und vermehren sich durch Zellteilung. Die meisten Biologen glauben heute, dass die Chloroplasten von Cyanobakterien abstammen. Man nennt diese Vorstellung Endosymbionten-Theorie, weil demnach die Chloroplasten und Mitochondrien als Endosymbionten in eukaryotischen Zellen leben.
Chloroplasten benötigen Licht für ihre Entwicklung und für die Fotosynthese. Für die Fotosynthese nutzen Pflanzen verschiedene Farbstoffe: polare Chlorophylle (blaugrünes Chlorophyll a und gelbgrünes Chlorophyll b) und Carotinoide (unpolare Carotine und polare Xanthophylle). Man sieht die Carotinoide erst im Herbst, wenn in den Blättern der meisten unserer Laubbäume in den Chloroplasten das Chlorophyll abgebaut wird.
Chloroplasten sind 5-10 µm große, kugelige oder linsenförmige Organellen. Die äußere Abgrenzung zum Zytoplasma bildet - ähnlich wie beim Zellkern, aber ohne Poren - eine Hülle aus einer äußeren und einer inneren Plastiden-Membran mit einem Spalt dazwischen. Die Grundsubstanz im Inneren heißt Matrix oder Stroma und enthält eigene Ribosomen sowie mehrere ringförmige DNA-Moleküle. Weil Chloroplasten Zucker produzieren, sind in ihnen Körnchen von Assimilationsstärke nachweisbar. Außerdem findet man im Stroma Lipidtröpfchen und große Stärketröpfchen, in denen die energiereichen Stoffe gespeichert werden, welche die Chloroplasten im Stroma produzieren. Die innere Plastiden-Membran ist an verschiedenen Stellen eingestülpt (leider zeigt unser Buch das nicht und beschreibt es widersprüchlich) und bildet im Inneren jedes Chloroplasten ein zusammenhängendes, dem endoplasmatischen Retikulum und dem Golgi-Apparat ähnelndes Membransystem, das viele kleine Hohlräume vom Stroma abgrenzt. Zusammen mit den sie umgebenden Membranen nennt man diese Hohlräume Thylakoide. Dabei unterscheidet man zwei Arten von Thylakoiden - die röhrenförmigen oder flächigen Stroma-Thylakoide und die aus runden Thylakoidstapeln bestehenden Grana oder Granathylakoide. Ein einzelner Stapel heißt Granum. Mehrere Stapel heißen Grana und werden durch Stroma-Thylakoide miteinander verbunden.
| Schema eines Chloroplasten | |
|---|---|
 | |
| Emmanuel Boutet, GNU-Lizenz für freie Dokumentation | |
| 1. äußere Membran, 2. Intermembranraum, 3. innere Membran, (1+2+3 = Hülle), 4. Stroma, 5. Thylakoidlumen (im Innern des Thylakoids), 6. Thylakoidmembran, 7. Granum (Granalamelle), 8. Thylakoid (Stromalamelle), 9. Stärke, 10. plastidäres Ribosom, 11. plastidäre DNA, 12. Plastoglobulus (kugelförmige Struktur aus Lipiden (Plural: Plastoglobuli) | |
Nur die Membranen der Thylakoide enthalten das Chlorophyll, das für die Fotosynthese gebraucht wird. Die Chlorophyll-Moleküle schwimmen zusammen mit Carotin-Molekülen und verschiedenen Proteinen und bilden mit diesen sogenannte Fotosysteme in einer Lipiddoppelschicht, die mit diesem Aufbau nicht untypisch für die Membranen von Zellen und Organellen ist.
Vakuolen sind mit Zellsaft gefüllte Hohlräume in Pflanzenzellen.
Der Tonoplast ist eine Membran und besteht daher aus einer Lipiddoppelschicht mit darin schwimmenden Proteinen. Er grenzt eine Vakuole vom Zytoplasma ab.
Direkt nach der letzten Zellteilung sieht die Zelle noch genauso aus wie eine embryonale Pflanzenzelle. Das Zytoplasma füllt noch gleichmäßig den gesamten Zellraum aus und in ihm schwimmen neben vielen anderen Organellen auch einige kleine Vakuolen. Während die Zelle wächst, werden die Vakuolen zahlreicher und größer, während das Cytoplasma kaum größer wird. Zwischen den Vakuolen bildet das Cytoplasma zunächst noch breite Schichten, später nur noch dünne Fäden und schließlich beschränkt es sich auf einen schmalen Rand zwischen dem Plasmalemma und einer vereinigten, großen Vakuole.
Der Zellsaft ist eine wässrige Lösung zahlreicher organischer und anorganischer Stoffe. Er befindet sich in den Vakuolen der Pflanzenzellen und besteht aus Reservestoffen und Stoffwechselendprodukten in wässriger Lösung.
Reservestoffe der Pflanzen sind Traubenzucker (Glucose), Fruchtzucker (Fructose) und Rohrzucker (Saccharose). Diese Stoffe werden vorübergehend aus dem Stoffwechsel heraus genommen und bei Bedarf wieder ins Zytoplasma zurück transportiert.
Stoffwechselendprodukte sind teilweise giftige Stoffe, die von der Zelle nicht mehr gebraucht, aber im Gegensatz zu Tieren von den Pflanzenzellen nicht ausgeschieden werden. Damit sie den Zellen nicht schaden, werden sie in der Vakuole endgelagert. Gerade weil sie teilweise giftig sind, schützen sie Pflanzen vor Tieren, die dadurch nicht allzu viel von einer giftigen Pflanze fressen können.
Pflanzen werden durch die Zellwand vor dem Platzen bewahrt. So können Pflanzenzellen durch ihre riesigen Vakuolen prall gefüllt und ähnlich stabil wie eine aufgepumpte Luftmatraze sein. Können die Vakuole aufgrund von Wassermangel nicht prall gefüllt werden, dann wird die Pflanze schlaff, sofern sie nicht durch Holz stabilisiert wird.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}